
В настоящее время в стране ведутся серьезные и многосторонние научные изыскания, посвященные диагностике и лечению нейроурологических заболеваний, разработаны и предложены современные алгоритмы по обеспечению ранней диагностики и патогенетически обоснованного лечения. Вместе с тем есть вопросы, нуждающиеся в специальном рассмотрении и обсуждении среди врачей узких
специальностей – урологов, неврологов, нейрохирургов. Это междисциплинарные аспекты реализации и нарушения функции нижних мочевыводящих путей. Одним из таких пограничных вопросов является возможное участие метасимпатической нервной системы в обеспечении физиологических функций мочевого пузыря и ее роль в формировании нейрогенных нарушений мочеиспускания [1–7].
В конце ХХ в. для обозначения комплекса вегетативных ганглиев и сплетений, расположенных в стенках полых мышечных органов, обеспечивающих их автономные свойства, российский физиолог академик А. Д. Ноздрачев предложил термин “метасимпатическая нервная система” и выделил ее в самостоятельный отдел вегетативной нервной системы. Метасимпатическая нервная система является периферической, эволюционно самой древней частью вегетативной системы. Сфера иннервации метасимпатической части автономной нервной системы ограничена и охватывает сугубо полые органы. Это иннервация базовая, все звенья ее рефлекторного пути локализуются только в интрамуральных ганглиях. Метасимпатическая часть автономной нервной системы не имеет своего центрального аппарата, и ее эфферентные связи с центральными структурами опосредованы нейронами симпатической и парасимпатической автономной нервной системы, образующими синаптические контакты на телах и отростках метасимпатических интернейронов и эффекторных нейронов. Эта автономия интрамуральных ганглиев, имея полный набор необходимых для самостоятельной рефлекторной деятельности звеньев – сенсорного, ассоциативного, эффекторного, с соответствующим медиаторным обеспечением представляет собой как бы собственный “мозг” органа. В эксперименте показано, что, обладая значительной независимостью от центрального и периферического регулирования, метасимпатическая нервная система способна обеспечивать адекватную рефлекторную деятельность органа при полной его денервации. Так, свежеэкстирпированный мочевой пузырь животного при достаточном наполнении через уретру теп-
лым солевым раствором способен к спонтанному опорожнению. При этом установлено, что пузырный автоматизм исчезает после рассечения интрамуральных ганглиев. В составе метасимпатической нервной системы имеются собственные сенсорные элементы, которые посылают в свои внутренние сети информацию о состоянии иннервируемого органа. Возможность самостоятельной работы вне влияния классических медиаторов (ацетилхолин, норадреналин) дает основание полагать наличие
других биологически активных веществ, инициирующих передачу импульса во внутренних сетях метастатической иннервации мочевого пузыря. С точки зрения органной принадлежности выделяют энтеро-, кардио-, уро-, везикулометасимпатическую нервную систему [8–10].
В процессе эволюции первоначально сформировавшаяся примитивная нервная система разделилась на анимальную и вегетативную нервную систему. Анимальная нервная система, связанная с деятельностью органов чувств и произвольной скелетной мускулатуры, обеспечивала приспособление организма к действию факторов окружающей среды. Ее функции контролируются сознанием. Вегетативная нервная система, регулируя деятельность внутренних органов, обеспечивала постоянство внутренней среды организма. В ответ на негативное влияние внешних факторов она, мобилизуя адаптационно-компенсаторные механизмы организма, способствовала выполнению функций анимальной нервной системы. При этом деятельность вегетативной нервной системы осуществлялась без участия сознания. Симпатическая часть автономной нервной системы брала на себя адаптацию организма к условиям внешней среды. Парасимпатическая часть автономной нервной системы способствовала сохранению постоянства внутренней среды организма. Метасимпатическая часть
автономной нервной системы обеспечивала врожденный автоматизм органа [11]. Вегетативная нервная система осуществляет управление работой органа посредством двух типов влияний – пусковых и корригирующих. Пусковые влияния используются в случае, если работа исполнительного органа не является постоянной, а возникает лишь с приходом к нему импульсов по волокнам автономной нервной системы. Если же орган обладает автоматизмом и его функция осуществляется непрерывно, автономная нервная система посредством своих влияний может усиливать или ослаблять его деятельность в зависимости от потребности. Это будут корригирующие влияния. Пусковые
влияния могут дополняться корригирующими [10–12].
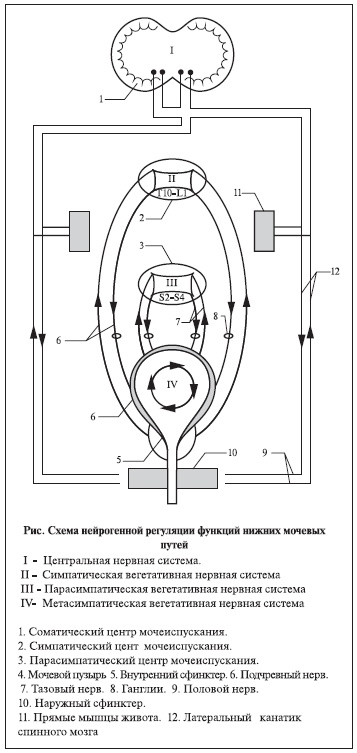
Рисунок. Схема нейрогенной регуляции функций нижних мочевых путей.
Урологам хорошо известно, что активация того или иного звена нервной системы определяет функциональный статус нижних мочевыводящих путей. В жизненном пространстве человека доминирует система удержания мочи, преимущественно регулируемая симпатическим отделом вегетативной нервной системы. Осознанное ощущение полноты мочевого пузыря опосредовано растяжением стенки органа возрастающим объемом мочи в фазу наполнения. При этом афферентные импульсы от рецепторов, расположенных в его стенке, по тазовому нерву поступают в крестцовый
отдел спинного мозга. Далее они направляются в центры мочеиспускания, расположенные в области моста и коры головного мозга (см. рисунок) [13–18]. Головной мозг снабжен органами внешнего контроля, которые оценивают сложившуюся жизненно важную ситуацию. Если в данный промежуток времени для конкретного индивидуума сложилась приемлемая обстановка, головной мозг, ощущающий позыв на мочеиспускание, конкретными действиями инициирует начало акта мочеиспускания. Одновременно плавно напрягаются мышцы живота, иннервируемые межреберными нервами, и расслабляются мышцы промежности за счет эфферентных соматических импульсов, достигающих мишени по срамному нерву. Это осознанный и управляемый этап мочеиспускания. Далее соматический импульс подавляет симпатическое доминирование над мочевым пузырем, обеспечивающим медленное накопление и хранение мочи, и активизирует парасимпатическое влияние на орган через эфферентные пути тазового нерва для быстрого и исчерпывающего опорожнения последнего [16, 19]. Отсутствие комфортных условий для акта мочеиспускания заставляет человека волевым решением подавить соматическую импульсацию от мочевого пузыря и передать команду симпатической иннервации продолжить процесс накопления мочи. Следующий позыв на мочеиспускание также может совпасть с отсутствием надлежащих условий. И вновь головной мозг подавляет реакции спинного мозга, направленные на избавление мочевого пузыря от нарастающего объема мочи. Позыв вновь перестает быть актуальным в поведении человека. Третий позыв на мочеиспускание тревожит головной мозг на пределе объемных возможностей мочевого пузыря. Условий для мочеиспускания по-прежнему нет. Сознание и воспитание не позволяют исполнить востребованный физиологический акт. Однако человек ощущает, что более не может сопротивляться нарастающему давлению мочи на управляемые мышцы промежности, уретры – и мощная струя как бы исподволь покидает мочевой тракт. Это результат повелительного позыва на мочеиспускание, который, игнорируя запретные усилия сознания и запрещающее координирующее влияние вегетативной нервной системы, побуждает автономную метасимпатическую нервную систему срочно и эффективно избавить мочевой пузырь от “угрожающего жизни” объема мочи. И только легкий румянец стыда укажет на вынужденное неповиновение мочевого пузыря центральной и вегетативной вертикали управления нервной системы. Принципиально важным является тот факт, что нормальное мочеиспускание возможно только при анатомической и функциональной сохранности мочевого пузыря и нервных структур, обеспечивающих этот процесс. Все остальные варианты опорожнения мочевого пузыря протекают в условиях дисфункции нижних мочевыводящих путей [20–27].
По результатам анализа современной научной литературы, посвященной этой проблеме, весь механизм накопления и опорожнения мочевого пузыря схематично нами представляется следующим образом. В процессе физиологического обеспечения работы нижних мочевыводящих путей организм человека создает и поддерживает определенный тонус поперечнополосатых мышц передней стенки
живота и промежности. В этих комфортных условиях ввиду наличия автономных свойств мочевой пузырь медленно накапливает мочу в расслабленный резервуар детрузора. Непроизвольное истечение поступившей на хранение мочи ограничивается повышенным тонусом внутреннего и наружного сфинктеров мочевого пузыря и исходным тонусом мышц промежности.
Физиологический тонус поперечнополосатых мышц тела человека указывает на адекватную работу головного мозга в рамках сознательного контроля за функцией нижних мочевыводящих путей в условиях адаптации организма человека к внешним факторам пребывания. Центральная нервная система одновременно оказывает корректирующее воздействие на работу вегетативной нервной системы, обеспечивающей гомеостаз и резервуарные функции мочевого пузыря. Физиологически преобладает симпатикотония мочевого пузыря. Детрузор расслаблен. Его размер медленно адаптируется под объем поступающей мочи. При этом ведущей функцией симпатической нервной системы является нивелирование внутрипузырного давления путем синхронного увеличения емкости мочевого пузыря. Парасимпатическая нервная система находится в состоянии покоя. Она не посылает импульсы на сокращение детрузора и расслабление внутреннего сфинктера. Все системы, регулирующие накопление и удержание мочи, пребывают в состоянии функционального равновесия.
Мочевой пузырь наполняется мочой до физиологически приемлемого уровня. Импульсы об этом поступают в центральные и периферические центры. Головной мозг с целью инициации акта мочеиспускания дает команду мышцам брюшного пресса на сокращение, а мышцам наружного сфинктера мочевого пузыря – на беспрепятственное обеспечение этого процесса. Эта импульсация оказывает пусковое воздействие на метасимпатическую часть нервной системы мочевого пузыря и корректирующее – на другие вегетативные центры. Симпатическое доминирование угасает, и нижние мочевыводящие пути переходят под влияние парасимпатической иннервации. Наступает фаза ваготонии мочевого пузыря. Детрузор сокращается, внутренний сфинктер мочевого пузыря расслабляется. Все происходит быстро, синхронно, и весь объем накопленной мочи покидает мочевой пузырь.
Головной мозг информируется органами внешнего контроля (слух, зрение, тактильные ощущения) о завершении акта мочеиспускания. Он дает команду на сокращение мышц промежности и расслабление передней брюшной стенки с последующим переводом их в режим физиологического тонуса. Одновременно автономные функции мочевого пузыря ставятся под охрану вегетативных центров, сопровождающих новый процесс наполнения мочевого пузыря в рамках обеспечения гомеостаза организма человека.
Как поведут себя мочевыводящие пути при потере доминирующего влияния головного мозга? Следует ожидать, что, потеряв связь с окружающим миром, врожденный автоматизм мочевого пузыря будет дезадаптирован относительно внешних факторов и попадет под покровительство вегетативной нервной системы. Работая преимущественно на сохранение внутреннего постоянства организма, вегетативная нервная система будет вынуждена компенсировать утрату центрального регулирования органа, но с явным акцентом на обеспечение внутренних потребностей.
Можно полагать, что целесообразная самостоятельная работа мочевого пузыря в рамках собственного
ритма будет подвергаться по сути корректирующему, но в реальности асинхронному вмешательству вегетативной нервной системы. Эта вполне определенная нейрогенная дисфункция органа классифицирована как гиперактивный мочевой пузырь. Клинически она проявляется повелительными позывами, при которых больной часто не в состоянии удерживать мочу. Важно отметить, что это заболевание, как правило, сопровождается полным опорожнением мочевого пузыря. По всей видимости, врожденный автоматизм органа, несмотря на некорректную импульсацию вететативной нервной системы, вполне состоятелен [6, 16].
Как отреагирует мочевой пузырь, если будет нарушена его симпатическая и парасимпатическая вегетативная регуляция? Известно, что при внезапном разрыве физиологических нервных связей в момент спинального шока имеет место угнетение всей рефлекторной деятельности спинного мозга. Акт мочеиспускания перестает быть произвольным. Исчезают спинальные рефлексы мочеиспускания, результатом чего становится острая задержка мочеиспускания [10, 27].
В процессе адаптации организма к факту разобщения форма нейрогенной дисфункции мочевого пузыря будет определена уровнем, степенью поражения спинного мозга, а также выраженностью феномена повышенной чувствительности нервной системы к медиаторам передачи импульсов, что является ее характерной посттравматической реакцией [11, 28]. Нейрогенная дисфункция мочевого пузыря при поражении спинного мозга выше места расположения спинальных симпатических вегетативных центров (Th10–L2), ответственных за иннервацию мочевого пузыря, клинически классифицируется как гиперрефлекторный нейрогенный мочевой пузырь с детрузорно-сфинктерной диссинергией. Этот вид нейрогенного нарушения мочеиспускания сопровождается постоянно повышенным внутрипузырным давлением и наличием остаточной мочи. Гиперрефлексия детрузора характеризуется автоматическим наполнением мочевого пузыря относительно небольшим объемом мочи и его внезапным опорожнением. Появление в мочевом пузыре остаточной мочи является следствием диссинергии детрузора и сфинктеров мочевого пузыря. [5, 7, 18, 29].
Поражение спинного мозга на уровне симпатических вегетативных центров (Th10–L2) ведет к параличу внутреннего сфинктера мочевого пузыря и повышению тонуса детрузора, в связи с чем возникает истинное недержание мочи. Мочевой пузырь практически пуст, так как, ощутив позыв, пациент не может удержать мочу [27, 29]. Если поражение локализуется выше сакрального парасимпатического центра мочеиспускания, но ниже грудного симпатического центра, гладкомышечный сфинктер мочевого пузыря сохраняет свою синергичность относительно функции детрузора и остаточной мочи нет. При поражении спинного мозга выше центров сегментарной иннервации мочевого пузыря (S2–S4, L1–L2) его рефлекторная активность осуществляется за счет самостоятельной деятельности спинномозговых центров. Чувство наполнения мочевого пузыря и позыв на мочеиспускание часто отсутствуют. Примитивный мочеиспускательный рефлекс замыкается в позвоночном канале на уровне S2–S4. Поскольку сам рефлекс не нарушен, вырабатывается автоматизм мочевого пузыря. По мере его растяжения от накопленной мочи раздражаются рецепторы растяжения, которые в свою очередь возбуждают парасимпатические нейроны, иннервирующие детрузор. Возникает неконтролируемое напряжение детрузора и расслабление внутреннего сфинктера. Мочевой пузырь опорожняется непроизвольно. Объем мочевого пузыря может быть небольшим, но обеспечиваемое детрузором давление становится достаточным для мочеиспускания [16]. При нарушении функции спинального рефлекторного центра (S2–S4) нейрогенное нарушение мочеиспускания классифицируется как гипоили арефлекторный мочевой пузырь. Детрузор не сокращается в ответ на заполнение мочевого пузыря, гладкомышечный сфинктер находится в сокращенном состоянии. Произвольный компонент акта мочеиспускания, регулируемый с помощью соматической иннервации (уровень S2–S4) и включающий управление наружным сфинктером, вспомогательными мышцами промежности, нарушен, и поперечнополосатый сфинктер не поддается произвольному контролю. Характерным проявлением данного вида нейрогенной дисфункции нижних мочевыводящих путей является наличие большого объема остаточной мочи в перерастянутом мочевом пузыре [27, 28].
Повреждение спинного мозга ниже уровня S2 и поражение периферической иннервации мочевого пузыря характеризуются сходной клинической картиной. Это арефлексия детрузора, несостоятельность внутреннего сфинктера и фиксированный тонус наружного сфинктера, не поддающийся произвольному расслаблению.
Восстановление пузырного рефлекса по окольным путям вегетативной нервной системы в позднем периоде после травмы при полном анатомическом перерыве спинного мозга – известный в клинике факт. По мнению одних авторов, это происходит за счет спинальной рефлекторной дуги. И нет основания говорить о формировании истинного автоматизма мочевого пузыря и классифицировать его как автономную форму мочевого пузыря [15].
По мнению других исследователей, в ряде случаев при сохранении функции интрамуральных ганглиев возможно восстановление рефлекторного опорожнения мочевого пузыря. Он начинает функционировать в “автономном” режиме, но не за счет спинальной рефлекторной дуги, а в связи с сохранением функции интрамуральных ганглиев [16, 27]. Немаловажным фактом является то, что клинические проявления и исход нейрогенных нарушений мочеиспускания зависят не только от уровня и степени поражения нервной системы, но и от длительности существования патологических условий для мочевого пузыря. Установлено, что сразу после возникновения нейрогенной дисфункции мочевого пузыря нарушается трофика тканевых структур органа. В его стенке развиваются нейродистрофические и воспалительные изменения. Длительное существование патологических процессов в мочевом пузыре усугубляет тяжесть вторичных дегенеративных изменений и ведет к склеротическому сморщиванию органа. Это требует скорейшей диагностики и патогенетически обоснованного лечения, поскольку бездействие может приводить к необратимым последствиям. Мочевой пузырь способен адекватно ответить на лечебные и реабилитационные мероприятия
только при условии сохранения в нем хотя бы части функционально полноценных элементов, что является важным прогностическим показателем в отношении возможности восстановления спонтанного мочеиспускания [6, 17].
Согласно рекомендациям Европейской ассоциации урологов по ведению больных с нейрогенным нарушением мочеиспускания, оптимальной лекарственной терапии нейрогенного нарушения мочеиспускания в настоящее время нет. Лучшим способом достижения максимальных результатов является комбинированная терапия. Из всех препаратов, доступных для лечения нейрогенного нарушения мочеиспускания, самыми эффективными признаны холиноблакторы, снижающие гиперактивность детрузора и улучшающие его резервуарные функции; α-адреноблокаторы, снижающие проявление дисрефлексии детрузора и улучшающие его эвакуаторные свойства; ботулотоксин, который при введении в детрузор снижает его гиперактивность и улучшает резервуарные функции, а при введении в наружный сфинктер снижает проявления дисрефлексии детрузора и улучает его эвакуаторные свойства (уровень доказательности 1, степень рекомендательности А) [30–38].
Арефлексия детрузора и несостоятельность внутреннего сфинктера, имеющие место при повреждении спинного мозга ниже уровня S2, подобны процессам, происходящим в экстирпированном в эксперименте мочевом пузыре животного с разрушенными интрамуральными ганглиями. Назначение в подобных клинических ситуациях холиномиметиков (парасимпатомиметиков) может вызывать
или усиливать сокращение детрузора, а комбинация их с α-адреноблакаторами – обеспечивать возобновление или улучшение мочеиспускания. Однако, учитывая частые и серьезные побочные эффекты данной комбинированной терапии, Европейская ассоциация урологов не поддерживает их применение [17, 36–39]. Вместе с тем сам факт обратимости блокады метасимпатической иннервации мочевого пузыря дает основание думать о химическом механизме ее происхождения и возможности медикаментозного устранения. Немаловажным является то, что все препараты с доказанной эффективностью при лечении нейрогенной дисфункции нижних мочевыводящих путей максимально
парализуют передачу нервных импульсов и приводят к временной вегетативной “денервации” органа. Можно полагать, что, ограждая мочевой пузырь от посттравматическогодезадаптирующего влияния вегетативной, а частично и центральной нервной системы, проводимая терапия создает условия для адекватного самостоятельного управления функции органа его метасимпатическим нервным аппаратом.
Допустим, что мочевой пузырь готов к автономному наполнению и опорожнению. Однако вместо ожидаемой автономности, которая вполне бы устроила пациента и врача, в клинической практике мы имеем гипер- или гипоактивность детрузора с диссинергией сфинктеров. Метасимпатическая нервная система демонстрирует свою неспособность в патологических условиях адекватно реализовывать накопительно-эвакуаторные свойства. Одной из объективных причин этого может быть дисфункция вставочного звена медиаторов, обеспечивающих перевод сигнала классических медиаторов (норадреналин, ацетилхолин) на собственные медиаторы метасимпатической сети. Это является серьезным поводом к поиску препаратов, вызывающих щадящее и избирательное разобщение дезадаптивного влияния центральной и периферической нервной системы на собственный, генетически детерминированный физиологический ритм метасимпатического обеспечения функции мочевого пузыря.
Существует ли физиологическая автономность мочевого пузыря? Вопрос риторический. Наверное, от взрослого человека трудно ожидать самостоятельного возобновления автономии этого органа ввиду наличия сложных и порой взаимоисключающих факторов, сопровождающих повреждение нервной системы и ее последствий. Вместе с тем надежда на то, что физиологический автоматизм при определенных условиях может быть реанимирован, вполне оправданна. Важно в самом раннем периоде появления нейрогенного нарушения мочеиспускания исключить или минимизировать факторы, ведущие к безвозвратной потере сократительных элементов детрузора. Это дает основания надеяться, что восстановление иннервации мочевого пузыря при определенных условиях пойдет по уже известному сценарию. Иными словами, перезагрузка генетически детерминированной физиологической программы нейрогенного обеспечения функции нижних мочевыводящих путей – вполне реальная для современной медицины задача.
Имея десятилетний опыт трансплантации трупной почки, мы всегда с восторгом наблюдали, как первая порция мочи появляется из дистального отдела мочеточника только что пересаженной почки, и мало обращали внимания на слаженную и вполне целесообразную перистальтику “чужого” мочеточника в новом теле, позволившую лицезреть этот удивительный и не до конца осознанный факт.



{kind=link}