Бульбоуретральные железы (БУЖ) являются малыми, или вспомогательными, железами мужской репродуктивной системы, к основной функции которых относится выделение щелочного секрета перед эякуляцией, что важно для нейтрализации кислой среды после прохождения мочи. Их дисфункция может приводить к различным заболеваниям, причем протекающим как бессимптомно, так и с нарушениями функционирования урогенитальной системы организма человека.
В представленной лекции отображены систематизированные данные о структурно-функциональной организации бульбоуретральных желез.
История открытия
БУЖ часто именуют «куперовыми» в честь английского врача и анатома Уильяма Купера (1666–1709), описавшего их в «Трактате о двух железах и их выводных протоках, недавно обнаруженных в человеческих телах» [1], опубликованном в 1699 г. В своей работе он не только изучил анатомическое строение и топографию данных органов, но и сделал верное предположение о выполняемой ими функции. Кроме того, он провел морфологическое сравнение БУЖ человека с подобными железами у крыс и свиней [2]. Первое же письменное упоминание об этом органе датируется 1684 г., когда во французском журнале «Journal des scavans» была опубликована небольшая заметка об исследованиях хирурга Жана Мери (1645–1722), в которой сообщалось об обнаружении им этих желез [3]. Однако о существовании БУЖ было известно и ранее. Так, по утверждению A. Riva et al., эти железы изображены на одной из таблиц итальянского анатома Джироламо Фабриция («Tabulae pictae», 1600), считавшихся длительное время утраченными и обнаруженных в 1609 г. в одной из библиотек Венеции [4].
Анатомия
БУЖ являются парными органами малого таза мужчины, имеющими приплюснутую овальную форму желтовато-бурого цвета и размером с небольшую фасолину (0,3–1 см в диаметре) [5]. Правая железа зачастую несколько крупнее левой, причем последняя нередко может отсутствовать [6]. Железы расположены книзу от предстательной железы в толще глубокой поперечной мышцы промежности, латеральнее и кзади от перепончатой части мочеиспускательного канала, друг от друга отстоят на расстоянии 0,6 см. Часть волокон упомянутой мышцы оказывает компрессионное действие на БУЖ, вследствие чего иногда называется m. compressor glandulae Cowperi. Кверху от них располагается луковично-губчатая мышца, оказывающая компрессионное воздействие на куперовы железы и стенки мочеиспускательного канала. В толще губчатые вещества луковицы полового члена иногда также встречаются добавочные БУЖ.
Железа состоит из долек, протоки которых сливаются в общий проток бульбоуретральной железы длиной 2–3 см, открывающийся в губчатую часть мочеиспускательного канала, несколько ниже его изгиба под лобковыми костями, вблизи срединной линии [5].
Кровоснабжение бульбоуретральных желез осуществляется артериями луковицы полового члена из внутренней половой артерии, а венозный отток – одноименными венами. Лимфатический отток происходит во внутренние подвздошные лимфатические узлы.
Симпатическая и парасимпатическая иннервация куперовых желез (равно как и других добавочных желез) осуществляется подчревным нервом и тазовыми внутренностными нервами соответственно. Чувствительная иннервация осуществляется ветвями полового нерва (крестцовое сплетение). Норадренергическая иннервация представлена только в слое гладкой мускулатуры, в железистом эпителии предположительно отсутствует. Стимуляция норадренергических волокон или введение катехоламинов приводит к сокращению мышц и выбросу секрета, однако не вызывает изменений в работе эпителия. Действуя через мускариновые холинорецепторы, парасимпатические волокна контролируют секреторную активность железистого компонента и, возможно, сокращение мышечных волокон стромы [7, 8]. Существует также информация о наличии пуринергической, нитрергической иннервации, а также об участии некоторых нейропептидов, таких как нейропептид Y (NPY), вазоактивный интестинальный пептид (VIP) [9, 10], кальцитонин-ген-связанный пептид (CGRP) и различные энкефалины [11, 12].
Сравнительная анатомия. БУЖ существуют практически у всех плацентарных животных и выполняют сходные функции. Однако у кротов, собак, ластоногих и китообразных они отсутствуют, а у человека развиты слабо. У надотряда афротерий (Afrotheria) они расположены дистальнее простаты, отличаются достаточно большими размерами. Важная особенность: у данной ветви наблюдаются сезонные отличия в размерах данных желез. Тем не менее у лавразиотерий (Laurasiatheria) среди животных отряда насекомоядных (Eulipotyphla, или Insectivora) куперовы железы не обнаружены: так, у многих видов либо присутствует исключительно простата, либо ее также сопровождают ампулярные железы. Среди отряда рукокрылых (Chiroptera) данные железы присутствуют практически у всех видов, за исключением нескольких, однако имеют небольшой размер и не столь выраженную секреторную функцию, в то время как среди представителей отряда парнокопытных БУЖ имеют все виды и наблюдается строгая сезонная зависимость их размеров, обусловленная изменением уровня андрогенов в крови. У животных отряда хищных (Carnivora), тупайи (Scandentia) и грызунов (Rodentia) строение и наличие БУЖ весьма вариабельны, однако все они окружены поперечнополосатой мускулатурой, что облегчает экскрецию и указывает на более активное функционирование данных желез. Среди добавочных желез сумчатых (Marsupials), в отличие от других отрядов, насчитывается три пары куперовых желез, также погруженных в ткани скелетной мускулатуры. Среди однопроходных (Monotremata) эти железы – и вовсе единственный тип добавочных половых желез, т.к. факт наличия у них простаты спорный [13, 14]. Функциональное сходство данных желез у различных видов подтверждается тем, что фосфолипаза A2 (PLA2), фермент, катализирующий специфический гидролиз эфирной связи в глицерофосфолипидах и играющий важную роль в акросомальной реакции [15], был выявлен в семенной плазме людей [16–18], быков, баранов [19] и мышей [20]. Более того, иммуногистохимическим методом выявлено наличие массы данного фермента как в эпителии предстательных желез мужчин, так и в бычьих семенных пузырьках и их секрете, в тканях БУЖ быка [21]. Это позволяет утверждать о сходстве структуры данного фермента у человека и некоторых других животных. Вероятнее всего, у других животных данный фермент также присутствует, но как минимум имеет другую конформацию, что осложняет его идентификацию иммуногистохимическими методами.
Эмбриогенез
Формирование БУЖ происходит параллельно с образованием предстательной железы и семенных пузырьков на 10-й неделе пренатального периода онтогенеза, под влиянием дигидротестостерона, а также в результате эпителио-мезенхимальных взаимодействий, как и в других железах. В частности, клетки мезенхимы, возникающие из урогенитального синуса, обладают андрогенными рецепторами и служат основной мишенью циркулирующих андрогенов, в то время как эпителиальные клетки, имеющие энтодермальное происхождение, не содержат рецепторов к андрогенам. В результате гормональной стимуляции дигидротестостероном мезенхимальные клетки начинают секретировать фактор роста фибробластов-10 (FGF-10) и трансформирующий фактор роста-β1(TGF-β1), что способствует выработке клетками энтодермы эпителия урогенитального синуса морфогенетического белка Shh (sonic hedgehog) [5]. Последний обеспечивает активацию формирования системы протоков, затем концевых секреторных отделов и ингибируется морфогенетическим белком BMP-4.
В основе всех этих молекулярных взаимодействий лежит регуляторное влияние транскрипционных факторов Hoxa-13 и Hoxd-13, управляющих органогенезом уретры, предстательной железы и БУЖ, в отличие от семенных пузырьков, развивающихся из компонентов семенного канатика и верхних отделов мезонефрального протока.
Процесс эпителиомезенхимальных взаимодействий продолжается при дифференцировке стромы и капсулы железы. Формирующийся эпителий стимулирует окружающую мезенхиму дифференцироваться в гладкомышечные клетки и соединительную ткань. При этом фиброэластическая капсула развивающихся желез содержит не только фибробласты и гладкомышечные клетки, но и скелетные мышечные волокна, возникшие в результате миграции миобластов сомитов для образования мочеполовой диафрагмы и соединившиеся с капсулой желез по причине регионального расположения промежуточной части урогенитального синуса, а впоследствии и перепончатой части уретры в этой области [22].
Железистые почки энтодермального происхождения врастают в окружающую мезенхиму из перепончатой части уретры в форме эпителиальных тяжей [22], растущих в заднем направлении параллельно развивающейся уретре, что приводит к дифференцировке протоков желез. Эпителий первоначально представлен однослойным кубическим, затем высоким призматическим, столбчатым, в концевых секреторных отделах в зрелой железе могут присутствовать все формы клеток от плоских, до столбчатых, а в главных протоках эпителий многорядный. Процесс морфогенеза протоков усиливается на 26-й неделе пренатального периода онтогенеза, что соответствует активному ветвлению протоков [22, 23].
Гистологическое строение
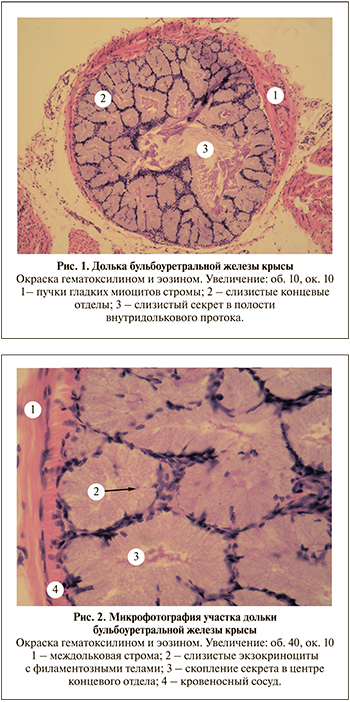 БУЖ представляют собой сложные трубчато-альвеолярные железы, окруженные капсулой из плотной соединительной ткани, от которой отходят септы, содержащие пучки гладких миоцитов, разделяющих орган на многочисленные дольки. Строма желез представлена рыхлой волокнистой неоформленной соединительной тканью, содержащей коллагеновые и эластические волокна, а также пучки гладких миоцитов (рис. 1). С вентральной стороны капсулы расположены группы скелетных исчерченных мышечных волокон [24].
БУЖ представляют собой сложные трубчато-альвеолярные железы, окруженные капсулой из плотной соединительной ткани, от которой отходят септы, содержащие пучки гладких миоцитов, разделяющих орган на многочисленные дольки. Строма желез представлена рыхлой волокнистой неоформленной соединительной тканью, содержащей коллагеновые и эластические волокна, а также пучки гладких миоцитов (рис. 1). С вентральной стороны капсулы расположены группы скелетных исчерченных мышечных волокон [24].
Концевые отделы образованы слизистыми экзокриноцитами, снаружи окружены миоэпителиоцитами, обеспечивающими экскрецию, однако их количество невелико. Миоэпителиальные клетки имеют веретеновидную или лентовидную форму, прикреплены к базальной мембране при помощи полудесмосом, а к секреторным клеткам – посредством десмосом. Их цитоплазма заполнена миофиламентами и большим количеством митохондрий, ядро уплощено. Расположены они близко друг к другу в достаточно хаотичном порядке, что затрудняет их визуализацию при помощи электронной микроскопии. Округлое ядро экзокриноцитов расположено в базальном отделе клетки, в апикальной части локализуются специфические гранулы, содержащие мукоидный секрет, а также ацидофильные тельца, имеющие веретеновидную форму. На морфологические параметры оказывает влияние их функциональное состояние [5].
Эпителий концевых секреторных отделов плоский, во внутридольковых отделах чаще выражен кубический или призматический однослойный, редко переходящий в многорядный. Концевые выводные протоки имеют однослойный многорядный эпителий. При избыточном накоплении секрета эпителиоциты уплощаются. Морфология клеток значительно варьируется в зависимости от стадии секреторного цикла, причем в одной локализации могут встречаться клетки как исключительно в начале или конце цикла, так и во всех промежуточных стадиях. На начальных стадиях секреции клетки имеют столбчатую форму, а секреторные гранулы неравномерно распределены в цитоплазме. При усилении продукции слизистого секрета количество секреторных гранул увеличивается и экзокриноцит удлиняется, приобретая пирамидальную форму. Секреторные гранулы могут отличаться в размерах и консистенции в зависимости от фазы их формирования. Экзокриноциты содержат капли мукоида и слизистые палочковидные включения – филаментозные тела (рис. 2) [25, 26].
Зрелые железистые клетки имеют пирамидальную форму, гладкую поверхность с редкими боковыми интердигитациями. При ультраструктурном анализе заметно, что ядра уплощены и располагаются у базального полюса клеток, имеется чередование эухроматина и конденсированного гетерохроматина. При изучении цитоплазмы электронно-микроскопическими методами обнаруживаются электронно-плотные гранулы со слизистым секретом, а также на периферии гранул обнаруживается плотное сферическое тельце.
В апикальной части клетки располагаются только секреторные гранулы, имеющие тонкую мембрану. Органоиды – комплекс Гольджи, митохондрии и гранулярная эндоплазматическая сеть – располагаются между мукоидными гранулами.
Помимо основных секреторных гранул также имеются нитевидные гранулы [25], или филаментозные тела (в соответствии с современной номенклатурой), ограниченные тонкой мембраной и содержащие филаменты, часто вокруг данных образований сосредоточены липидные капли. Иногда эти два типа гранул могут объединяться: такие гранулы расположены базально, и после слияния нитевидный компонент исчезает. Важно отметить, что экзоцитоза нитевидных гранул не наблюдается. Вероятнее всего, секреция их содержимого происходит только в комплексе с мукоидными гранулами, по аналогии с клетками слизистых носа и слюнных желез [26].
Клетки, находящиеся на начальной стадии секреции, малочисленны, отличаются высоким содержанием светлого хроматина и хорошо очерченным ядрышком. Ядро обычно сферической, реже – яйцевидной формы, однако иногда наблюдаются глубокие инвагинации. В перинуклеарной области располагается аппарат Гольджи. Эндоплазматическая сеть умеренно развита, ее короткие цистерны локализуются в базальной части клетки и окружены митохондриями. В цитоплазме наблюдается масса свободных рибосом, агрегированных в так называемые розетки. Секреторные гранулы малочисленны, их размер несколько меньше, чем у таковых в зрелой клетке. Филаментозные тела также малочисленны и по большей части находятся над ядром [25].
В пирамидальных клетках секреторные продукты высвобождаются в просвет по апокриновому (у человека) или мерокриновому (у млекопитающих) механизму.
Выводные протоки БУЖ покрыты многорядным столбчатым эпителием, причем переход от секреторного эпителия происходит достаточно плавно, вследствие чего в эпителии протоков сохраняются одиночные экзокриноциты или даже их скопления [26]. Форма эпителиоцитов протоков неправильная, периодически имеет место соединение нескольких протоков в один; таким образом, секрет из долек может поступать в уретру либо напрямую, либо через общий проток. Апикальная поверхность клеток покрыта микроворсинками и может обладать инвагинациями, иногда образует выпячивания в просвет протока [25].
В цитоплазме могут находиться электронно-плотные гранулы, тем не менее не участвующие в механизме экзоцитоза. Ранее считалось, что они могут выделять серозный секрет, однако слаборазвитый синтетический аппарат (гранулярная эндоплазматическая сеть и аппарат Гольджи) указывает на ошибочность данной точки зрения.
Паренхима органа претерпевает изменения в ходе онтогенеза, ее относительная площадь у грудных детей минимальна, максимальных значений она достигает к концу пубертатного периода (в среднем к 16 годам). Помимо площади изменяется и соотношение компонентов: доля соединительной ткани и мышечных волокон постепенно снижается и достигает минимального значения в период полового созревания [27].
Физиология
Экскреторная деятельность. Куперовы железы принимают участие в акте эякуляции. Их щелочной секрет представлен вязкой прозрачной субстанцией, богатой различными гликозаминогликанами, такими как галактозамин, сиалопротеин, метилпентоза, галактуроновая кислота [28].
Цикл сексуальной реакции мужчины состоит из четырех фаз: возбуждение, фаза плато, оргазм и заключающая фаза [29]. Выделение секрета происходит во время возбуждения и продолжается в фазе плато, что выполняет ряд задач. Объем жидкости, именуемой преэякулятом, колеблется в границах 0,2–5 мл в зависимости от уровня сексуального напряжения в фазу плато [24], но обычно составляет 1% от семенной жидкости. Щелочной характер субстанции способствует нейтрализации кислых остатков мочи в мочеиспускательном канале мужчины непосредственно перед эякуляцией [24]. Секрет данных желез выполняет функции лубриканта и попутно снижает кислотность влагалища [5, 30, 31].
Экспериментально подтвержден факт, что секрет не содержит сперматозоиды, т.к. в выделениях этой железы до эякуляции сперматозоиды обнаружены не были, в то время как в эякуляте они имели место быть [29].
Доказано, что поражение бульбоуретральных желез оказывает неблагоприятное воздействие на фертильность сперматозоидов. Экспериментально установлено, что при сирингоцеле БУЖ самцы мышей теряли фертильность. Тем не менее исследований, направленных на выявление зависимости репродуктивной функции мужчин от патологических процессов в куперовых железах, до сих пор проведено не было. Предположительно это обусловлено скрытым течением заболеваний и несовершенством диагностики [32, 33].
Иммуногистохимическим путем было выявлено, что БУЖ секретируют муцин MG1 [34], ранее считавшийся исключительно компонентом слюны. Секреции MG2 не отмечалось, хотя не исключено, что значения его концентрации ниже порога чувствительности метода [34]. Предполагается, что, как и другие муцины, MG1 играет вспомогательную роль для транспорта спермы по мужскому и женскому половому тракту [35], создает плотную тонкую защитную пленку на внутренней поверхности уретры, прочно адгезирующую с поверхностью эпителия [34]. Несмотря на то что ранние исследования отмечают ингибирующее действие MG1 и MG2 на HIV-1 и подтверждают защитные свойства муцинов против ВИЧ в слюне [36], в жидкостях репродуктивной системы инфицированного партнера концентрация вируса значительно больше, что делает MG1 неспособным предотвратить заражение мужчины [37].
Среди всех добавочных желез мужской репродуктивной системы БУЖ выделяют наибольшее количество плазминогена, стимулирующего фибринолитическую активность эпителия репродуктивного тракта [38]. Предполагается, что это необходимо для лизиса фибриновых отложений [39] и поддержания семенной жидкости в жидком состоянии [40]. Возможно, это также играет роль в капацитации сперматозоидов и акросомальной реакции.
Эндокринная функция и гормональная регуляция. Иммуногистохимическим методом выявлено, что наряду с клетками других органов репродуктивной системы клетки стромы куперовых желез уже в фетальном периоде экспрессируют стероидную 5α-редуктазу II типа [41]. Доказано, что данный фермент служит как аутокринным, так и паракринным медиатором роста и дифференцировки мужской репродуктивной системы, является ключевым для нормального развития простаты и наружных половых органов по мужскому типу, так как катализирует превращение тестостерона в более сильный по действию гормон – дигидротестостерон [41, 42]. Мутации гена или иные причины, вызывающие дефицит 5α-редуктазы II типа, могут приводить к таким нарушениям, как псевдогермафродитизм и псевдовагинальная перинеоскротальная гипоспадия [43, 44].
Куперовы железы очень чувствительны к изменениям гормонального фона, особенно к уровню половых гормонов. В своем исследовании R.E. Heller сравнивал гистологическое строение бульбоуретральных желез здоровых и кастрированных крыс. После удаления яичек наблюдается инволюция куперовых желез, характеризующаяся на 10–20-й день резким снижением количества ацинусов и уменьшением высоты железистых эпителиоцитов на 30%.
К 25–30-му дню значительно возрастает доля соединительной ткани в ацинусах и возникает атрофия крупных выводных протоков. Спустя 60 дней общий объем и размеры железы значительно уменьшаются. Ключевым моментом для данного периода является снижение вплоть до исчезновения ряда цитоплазматических включений и секреторных везикул. Роль половых гормонов в данных процессах доказывает тот факт, что при введении дозы половых гормонов сразу после кастрации удавалось не только сохранить железы в исходном состоянии, но и даже практически полностью восстановить исходный размер и секреторную активность спустя 190 дней. Соответственно, избыток гормонов приводил к гипертрофии желез [45].
Защитная функция. Полагается, что куперовы железы принимают участие в иммунной защите мочеполового тракта, выделяя массу гликопротеинов, в том числе простат-специфический антиген (PSA) и просапозин (PSAP). Иммуногистохимические исследования трупных образцов после цистопростатоуретрэктомии показали, что, хотя PSA и PSAP в основном вырабатываются тканями предстательной железы, это свойственно и для бульбоуретральных желез [46].
Около 10% от популяции слизистых клеток БУЖ составляют Т-лимфоциты. Большинство из них цитотоксические (CD8+) и составляет в среднем 65,7% от общего числа Т-лимфоцитов (CD3+) эпителия железы. Субпопуляция CD4+-лимфоцитов оказывается меньшей, при этом эти клетки располагаются преимущественно в интерстициальной соединительной ткани. Субпопуляции моноцитов и макрофагов скудны и представлены большей частью в собственной пластинке [47].
В численности практически всех субпопуляций лимфоцитов БУЖ уступают предстательной железе. В этом они очень сходны с семенными пузырьками. Это объясняется тем, что хорошо представленный иммунный барьер необходим лишь в случае абсорбции антигенов семенной жидкости, что в данных железах происходит редко [48]. При этом относительное количество Т-супрессоров в тканях куперовых желез оказывается также меньшим (в чем также сходство с семенными пузырьками), чем в тканях предстательной [49].
Переменное, но все же малое количество В-лимфоцитов (локализующихся преимущественно в интерстициальной соединительной ткани) по сравнению с численностью таковых в простате также говорит о преимущественной роли предстательной железы в локальной секреции иммуноглобулинов и иммунной защите генитального тракта [47, 50].
Присутствие макрофагов может говорить о дополнительной функции иммунного барьера БУЖ. Данные клетки могут участвовать в клеточном и гуморальном ответе, выполняя фагоцитарную и антиген-презентирующую функцию. Гистохимическое окрашивание показывает, что большая часть макрофагов активны и секретируют HLA-DR-антигены. Однако меньшее, чем в тканях предстательной железы, количество макрофагов, моноцитов и их антител с большой вероятностью подтверждает различные роли желез в иммунной защите тракта [47].
Что касается противоопухолевой защиты, клетки бульбоуретральных желез активно экспрессируют онкосупрессорный белок p53, выполняющий антиканцерогенную функцию. Это объясняет крайне низкую частоту малигнизации, в частности, по сравнению с опухолями предстательной железы [33].
Клинические аспекты
Различные заболевания бульбоуретральных желез, как правило, не приводят к возникновению серьезных осложнений, и их обнаружение по большей части случайно при рентгенологическом исследовании. Из-за отсутствия симптомов истинная частота возникновения патологий неизвестна, однако, согласно статистике, они встречаются у 2–3% мужчин [23, 32]. Считается, что среди взрослых мужчин большинство случаев возникают либо вследствие травмы, либо в виде осложнений инфекционной болезни мочеполовых путей, в то время как у детей основной причиной служат врожденные аномалии, такие как облитерация протока или возникновение кист [51]. Редким смертельным осложнением является внутриутробная обструкция мочевыводящих путей, вызванная кистой куперовой железы значительных размеров.
Самые распространенные заболеваниям БУЖ – это куперит (воспаление железы) и сирингоцеле (syrinx – свищ, cele – отек).
Основной причиной возникновения куперита является инфекционное поражение железы, сопровождающееся классической картиной воспаления. Она в свою очередь трудно отличима от таковой при медленнотекущем уретрите или простатите, что значительно затрудняет диагностику [32]. К наиболее распространенным возбудителям относятся E. faecalis, E. faecium, E. coli, K. pneumonia, E. aerogenes [52]. К серьезным осложнениям куперита относят абсцесс, однако вероятность его возникновения крайне мала. Так, в работе M. Valentino et al. указывается, что случай возникновения абсцесса куперовой железы наблюдался лишь единожды на 19 тыс. за период исследований длительностью 23 года [53]. За всю историю наблюдений абсцесс БУЖ регистрировался всего 9 раз, 7 из которых были следствием перенесенной гонококковой инфекции [32].
Сирингоцеле представляет собой кистозное расширение выводных протоков БУЖ. Существует несколько классификаций данной патологии. Так, согласно M. Maizles et al. в зависимости от результатов рентгенологических и эндоскопических исследований и от степени увеличения диаметра протока выделяют четыре типа: простой, кистозный, перфорированный и разорванный [54]. К простому типу относят небольшой отек дистальной части протока, к кистозному – закупорку протока, которая приводит к расширению его дистальной части, что в свою очередь может оказывать давление на бульбарную часть уретры, как следствие – стеноз и расширение ее проксимальной части. При перфорированном типе расширение проксимальной части уретры выглядит как дивертикул бульбарной части уретры, причем зачастую отмечается помутнение куперовых желез. Разорванный тип характеризуется расширением бульбарной части протока при его изолировании от проксимальной части вследствие закупорки [6]. Вязкая консистенция вырабатываемого секрета наряду с достаточно большой длиной выводящих протоков может приводить к их обструкции, что симптоматически проявляется в виде болей в тазу, особенно при мочеиспускании [52].
В то же время другие специалисты [55] различают два типа – открытый и закрытый, подразделяющиеся в зависимости от наличия (или отсутствия) сообщения с уретрой. Открытый тип чаще встречается у взрослых и проявляется клинически, в то время как закрытый тип в большинстве случаев – врожденная аномалия либо следствие инфекции и по большей части протекает бессимптомно. К основным проявлениям открытого сирингоцеле относят гематурию, вторичную инфекцию мочевыводящих путей, патологические выделения из уретры и выделения после мочеиспускания; в то время как к закрытому – дизурию, задержку мочеиспускания и боль в промежности.
Бульбоуретральные, или куперовы, железы представлены добавочными железами мужской урогенитальной системы широкого круга плацентарных животных. К отрядам, имеющим данные железы, относятся Chiroptera, Carnivora, Scandentia, Rodentia, Marsupials, Monotremata, Primates и др. Бульбоуретральные железы имеются также у человека, хотя и не достигают высокой степени развитости. Функциональное и структурное сходство изучаемых желез у представителей разных видов подтверждается иммуногистохимическими методами (таким образом, PLA2 сходной структуры выявлена у людей, быков, баранов и мышей).
Прежде всего куперовы железы – это сложные трубчато-альвеолярные железы, развивающиеся одновременно с простатой и семенными пузырьками урогенитального синуса. Секреторная деятельность БУЖ тесно связана с актом эякуляции, при этом обеспечивает нейтрализацию кислых остатков мочи и более низкой pH женского влагалища, снижение трения во время полового акта. Более того, есть причины предполагать антимикробную и фибринолитическую активность секрета БУЖ. Данные железы служат иммунным барьером для урогенитальных инфекций, а также в период роста организма синтезируют важные аутокринные и паракринные факторы развития мужской мочеполовой системы.
Что касается заболеваний куперовых желез, наиболее распространены инфекционные купериты и сирингоцеле. Обычно они не приводят к серьезным осложнениям и практически не поддаются диагностике, вследствие чего оказывается достаточно тяжело оценить частоту встречаемости и значения патологий бульбоуретральных желез.
